櫻井 武
大脳ニューロンはそれぞれが固有に発火しているが、ヒトは眠るとまずノンレム睡眠に入り、ノンレム睡眠のときは神経細胞の発火が低下してだんだんオフ期とオン期のタイミングがそろい、スポーツイベントのウェーブのように同期するようになる。90分ほどたつとまた活動をはじめ、ばらばらに固有の発火をみせるようになる。これがレム睡眠で脳は活動時と同じかそれ以上に強く活動しているが、感覚系は視床のレベルで 運動系は脊髄のレベルで情報伝達が遮断されておりオフラインの状態である。レムの中断でレム潜時(レムがおこるまでのノンレムの長さ)の短縮が起こる。つまりレム睡眠は深い睡眠であるノンレム睡眠とは別に、レム睡眠として守られる理由がある。
以下の二つの記憶は成立のメカニズムに差があるが、睡眠はどちらにも大きな効果
宣言的記憶 一般的な自己に関する記憶 エピソード、意味、空間記憶
非宣言的記憶 手続き記憶(大脳皮質のほかに小脳や大脳基底核が関与)、情動睡眠を減らすと長期増強(情報を送る側のニューロンを高頻度で刺激するとそのシナプス結合が強化される現象)が減弱する。椎体細胞の周期的発火がニューロンや細胞間のつながり再構築に必要なのかも 配線のつなぎ変えを常時行っているが、電源入れっぱなしでメンテナンスをするわけにはいかない。いったんスリープモードが必要なのでは。
大脳皮質6層 140億の神経細胞 シナプスの伝導効率は常に変化 数や構造も刻刻変化 細胞膜にはイオンチャンネルというたんぱく質やトランスポーター(特定物質を取り込む)、受容体(特定物質をつかんでシグナルと細胞内に伝える)がある
脳表近くの樹状突起の電気的な変化(シナプス後電極)の集合が脳波 脳波の信号の強さはニューロンの活動がどれだけ同期するかによる
レム期は覚醒時より皮質の活動は増加 脳幹から脊髄に運動ニューロンを麻痺させる信号が出て、全身の骨格筋は麻痺、脳の命令は筋には伝わらない。
オンラインは覚醒 スリープモードはノンレム オフラインはレム
ノンレムの血流 視索前野の睡眠中枢以外は低下
覚醒時によく使う、左側、前頭葉は特に活動低下
レム睡眠は橋被蓋に中枢があり、アセチルコリン作動性ニューロンが活動、海馬や扁桃体などの大脳辺縁系も活動増加。視覚野へも信号。前頭前野背外側部の機能低下により、自己を客観的にとらえるメタ認知が欠如し、奇妙な夢、一時視覚野は休止しているが、高次視覚野である視覚連合野は活発に活動し、視覚イメージが作られる。
覚醒β→閉眼α→ノンレムθ→ノンレムδ→レムでは覚醒時に似た低振幅速波

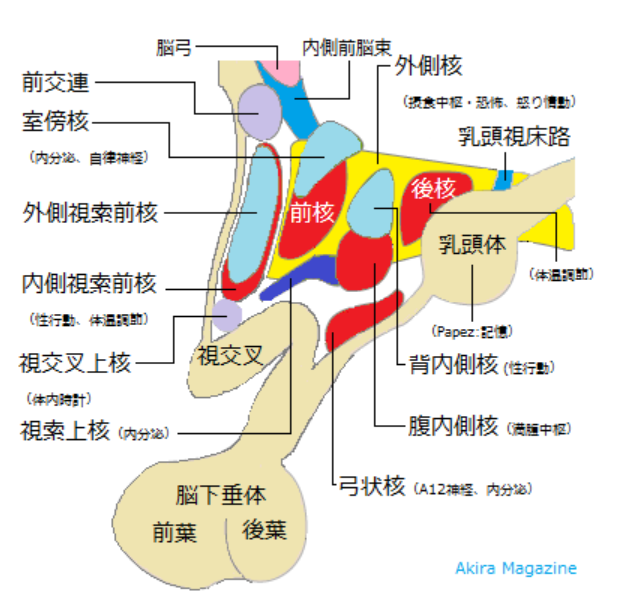
視床下部は
①自律神経、恒常性の中枢
②情動や本能活動ともかかわり、
③睡眠と覚醒のコントロール中枢でもある。
すぐ隣に脳幹があり、ここを通じて大脳皮質のモードを変える。上行性脳幹網様体賦活系により覚醒とレム睡眠の両方が起こる。GABA、グルタミン酸などの樹状突起に作られた棘突起を介するシナプスで局所的に働く通常の神経伝達(Eメール)のみではなくモノアミン作動性システムとコリン作動性システムが関与。
①ノルアドレナリン青斑核、セロトニン縫線核、ヒスタミン結節乳頭体から広範投射系とよばれる軸索を伸ばす 軸索末端は数珠上に膨らみ、モノアミンを分泌して周辺の多数のニューロンに影響を与える(館内放送)
②コリン作動性 橋の外背側被蓋核と脚橋被蓋核は視床に投射しここを介して脳全体に影響
覚醒時①②とも↑ ノンレム①②とも↓ レム①↓②↑
①は体温調整にも関与、レムではできない 山で眠ると凍え死ぬ
覚醒と睡眠はシーソーで睡眠よりである オレキシンはこの覚醒の重しとなり、覚醒相の安定化につながる
視交叉上核 の体内時計の支配をうけて松果体からメラトニン分泌
睡眠のタイミングは脳内のアデノシンと睡眠負債のバランス。アデノシンは視索前野のGABA作動性ニューロンを刺激し、ここから①②を抑制して、睡眠が起こる。
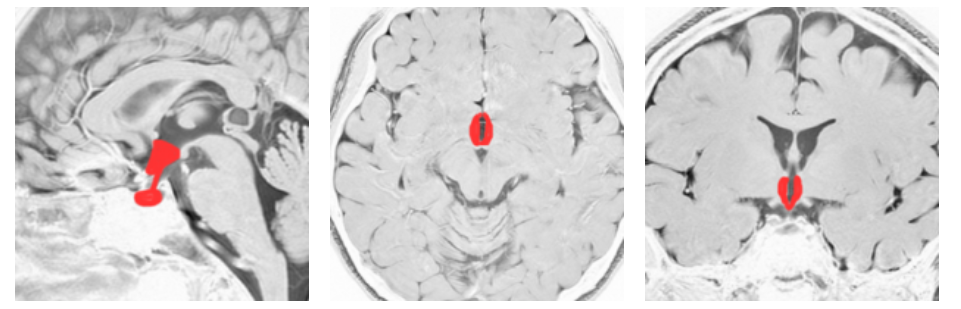
オレキシンは 摂食中枢と考えられてきた 視床下部外側野のニューロンにより産生され、このニューロンはモノアミン作動性ニューロンに軸索を伸ばしている。 オレキシンの欠損でナルコレプシーが起こる 。突然の強い眠気と情動脱力発作と金縛り(睡眠麻痺) 寝入ってすぐにレム期に入ってしまい、通常とちがって、前頭前野背外側部は活動低下しておらず、金縛りを実感できる。視床下部腹内側部は満腹中枢。視床下部前部には 視索前野があり、ここが睡眠を作り出す睡眠中枢。
注意には前脳基底部のコリン作動性ニューロンが大きな役割、脳幹のコリン作動性ニューロンとは別集団。オレキシン作動性ニューロンとこのコリン作動性ニューロンは相互に刺激、注意の発動で覚醒も高める。オレキシン作動性ニューロンへの入力は 情動。扁桃体から直接、間接に多くの入力を受ける 情動は、オレキシンニューロンを介して覚醒かつ、交感神経刺激。原因がはっきりした心配でなく意識には不安や心配がのぼっていなくても脳はそうした用意を感じ続けることがある。ストレスが慢性的にかかると情動反応は起こっていても、海馬のシステムに変調をきたすため記憶が曖昧になって、何が不安や心配の原因になっているのか本人が実感できなくなる。これが不安性障害で、不眠を伴うことが多い。意識は自分に起こっていることをすべて把握しているわけではない。オレキシン作動性ニューロンは全身の栄養状態をモニターすることができる。
視床下部の視索前野と腹外側視索前野VLPOは睡眠時にのみ活動するGABA作動性ニューロンが存在。この睡眠ニューロンは覚醒のモノアミンニューロンを抑制かつオレキシンニューロンを抑制 モノアミンニューロンは睡眠ニューロン、オレキシンニューロンを抑制 オレキシンはモノアミンを活性化 この自分を抑制するものを活性化する仕組みがネガティブフィードバック(活性化するものを抑制する場合も)という モノアミンが抑制され、眠くなってくると、オレキシンへの抑制も減り、 ネガティブフィードバック でモノアミンをあげようとして覚醒が維持される。カフェインはVLPOの睡眠ニューロンに発現するアデノシン受容体A2A受容体に拮抗的に働く
モノアミン アミノ酸からカルボキシル基をとった構造物 ヒスタミンはアミノ基が二個あるので正確にはモノアミンではないが、便宜上含める。ドパミン報酬系、ノルアドレナリン精神興奮はカテコールアミン、セロトニン安心はインドールアミン。統合失調症にはドパミンの拮抗が必要で、セロトニンやノルアドレナリンはうつと関係
脂肪レプチン 視床下部 食欲抑制 オレキシンニューロンを抑制
胃グレリン 食欲増進 オレキシンニューロンを興奮
オレキシンニューロンは栄養状態に応じた摂食行動を維持するために、覚醒を制御する。血糖が下がると発火、空腹ではオレキシンを開始覚醒レベルが上がる。眠れない。
覚せい剤はモノアミントランポーターを抑制し、神経伝達物質がいつまでも細胞間隙にいるようにする
睡眠導入剤 BZはGABAニューロンを介して脳全体を抑え込む 非BZはGABA受容体のサブタイプに選択制がある
視覚野 左側からの視野は右脳に 右側からの視野は左脳に 左目の視野が右に入るのではないよ念のため
レム睡眠行動異常の原因は不明。コリン作動性ニューロンが脊髄以下をブロック。
